
Dans les derniers billets de cette série, nous avons introduit l’idée que des caractéristiques individuelles (comme les séquences d’un gène individuel) peuvent ne pas toujours correspondre à la phylogénie, ou arbre généalogique des espèces pour un groupe d’organismes apparentés. Le tri de lignéess incomplet en est une cause, des similitudes qui surviennent par des événements indépendants en sont une autre. De telles caractéristiques auraient l’apparence superficielle d’avoir été héritées d’un ancêtre commun, mais seraient en fait des exemples d’homoplasie : des caractéristiques partagées entre des espèces qui n’ont pas été héritées d’un ancêtre commun.
Oiseaux et plumes
Le vol actif chez les oiseaux et certains mammifères (comme les chauves-souris) est un exemple classique d’homoplasie. L’arbre généalogique des espèces des oiseaux, des chauves-souris et des mammifères qui ne volent pas (par exemple les souris) nous montre que toutes les espèces de mammifères sont bien plus proches entre elles qu’avec les oiseaux. Afin d’expliquer la caractéristique du vol actif partagée par les chauves-souris et les oiseaux, il faut donc recourir au modèle de l’homoplasie, et les comprendre comme des événements indépendants qui surviennent sur deux lignages séparés :
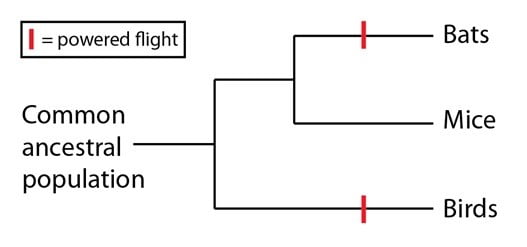
Population ancestrale commune, Bats= chauve souris, Mice=souris, bird=oiseau
L’autre explication possible, que le vol actif est homologue entre les chauves-souris et les oiseaux (et est ainsi présent chez leur dernier ancêtre commun), supposerait que tous les mammifères sauf les chauves-souris auraient perdu cette capacité (pour ne rien dire des séquences ADN qui soutiennent l’arbre généalogique des espèces ci-dessus). Au-delà de cette évidence, il y a aussi de bonnes raisons de penser que le vol actif est survenu indépendamment chez les chauves-souris et les oiseaux en comparant leur anatomie. Les oiseaux utilisent des plumes attachées tout au long de leurs membres supérieurs pour décoller. Par contraste, les chauves-souris utilisent une membrane pour former leurs ailes, et cette membrane est attachée entre leurs doigts ainsi qu’à leur corps :

Les deux solutions fonctionnent bien, mais lorsque nous divisons la caractéristique étendue de « vol actif » en ses éléments, nous voyons que si ce caractère dans son ensemble est convergent, les éléments sous-jacents ne le sont pas. Cette observation soutient que le vol actif des oiseaux et des mammifères est survenu séparément.
Homoplasie et homologie
Comment une simple homoplasie d’une séquence ADN peut survenir en utilisant une phylogénie peut être illustré. Supposons que trois espèces ont la séquence suivante pour une portion du même gène :

En se basant sur ces seules données, la phylogénie la plus simple (la plus parcimonieuse) serait comme suit :
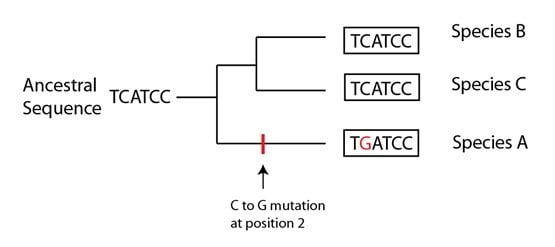
En se basant sur ces données, on inférerait que la séquence ancestrale est “TCATCC” et la branche de la phylogénie qui conduirait à l’espèce A aurait eu une mutation pour expliquer la différence au niveau de la séquence. En l’absence d’autres indices, cette phylogénie serait la plus appropriée pour ces données.
Cette image bien ordonnée, cependant, pourrait être dérangée par d’autres données, des données qui démontrent que le simple arbre généalogique des espèces que nous avons dessiné ci-dessus est en fait inexact. Dans ce cas, nous devons ordonner les séquences ci-dessus dans un arbre généalogique des espèces différent, ce qui signifie que nous devrons expliquer le patron en utilisant plus qu’un événement de mutation. Examinons un exemple hypothétique pour en montrer le processus.
Supposons que les données des séquences de plusieurs centaines de gènes additionnels sont comparées pour ces trois espèces ainsi que pour un certain nombre d’autres espèces apparentées qu’on ne montre pas sur notre arbre. Supposons aussi que ces données soutiennent fortement un autre arbre que celui que nous avons généré : dans la vaste majorité des cas, les données soutiennent un arbre sur lequel les espèces A et B sont apparentées de près, et l’espèce C apparentée de bien plus loin. Cela nous « forcerait » à redessiner l’arbre généalogique des espèces comme suit, en plaçant nos courtes séquences en un patron différent aux côtés de leurs espèces :
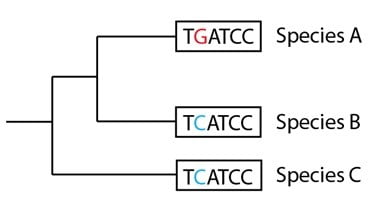
Supposons également que les données de la séquence ADN pour la séquence de ce gène particulier indiquent avec les espèces additionnelles qui ne sont pas montrées sur notre arbre généalogique des espèces que la séquence ancestrale avait un « T » en deuxième position et non un « C » :
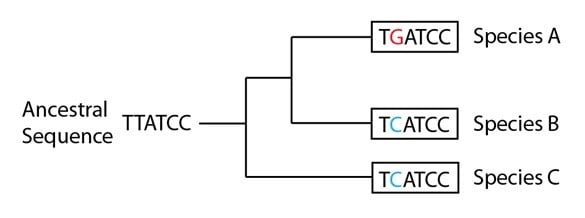
Il nous faut maintenant rendre compte pour les trois espèces de notre arbre du fait qu’elles n’ont pas une séquence ancestrale en seconde position, ainsi que comprendre les événements mutationnels qui ont conduit au patron que nous voyons ici. Vous noterez que nous sommes toujours contraints de proposer l’explication la plus parcimonieuse pour l’ensemble de ces données, mais pour ce gène particulier, nous sommes forcés d’invoquer des événements mutationnels multiples pour correspondre au patron de l’arbre généalogique des espèces. Nous faisons ce choix, cependant, parce que ce serait encore plus improbable que des événements mutationnels aient formé le patron de centaines d’autres séquences de gène de façon coordonnée ; et ces autres séquences soutiennent cette version de l’arbre généalogique des espèces.
Si vous essayez de « résoudre » l’arbre généalogique génétique en ajoutant des événements mutationnels à l’arbre généalogique des espèces, vous vous rendrez vite compte qu’au moins trois événements mutationnels sont nécessaires pour produire le patron observé. D’autres solutions utilisent plus de trois événements mutationnels, mais elles sont des explications moins probables. L’une des solutions possibles est exposée ci-dessous :
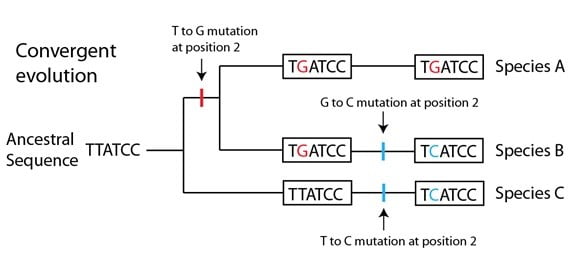
Sur la branche de la phylogénie qui conduit aux espèces A et B, une mutation (T à G) précède la divergence des espèces A / B (représentée par la barre rouge). Une seconde mutation a lieu ensuite sur le lignage qui conduit à l’espèce B, et change le G en même position en C (représentée par une barre bleue). Indépendamment de cela, le lignage qui conduit à l’espèce C a aussi une mutation sur cette position, qui change le T ancestral en C (représentée aussi par une barre bleue). Le résultat final est que deux séquences (dans les espèces B et C) sont devenues identiques, alors qu’aucune n’a hérité du « C » en seconde position de leur ancêtre commun. En d’autres termes, elles sont arrivées à la même « destination » à partir de points de départs différents, ou ont « convergé » en une séquence commune. On appelle ce phénomène évolution convergente. Pour ces deux espèces, le « C » en seconde position n’est pas homologue (une similitude héritée d’un ancêtre commun) mais plutôt une homoplasie, c’est-à-dire une similitude qui résulte d’événements indépendants sur deux lignages.
Une homoplasie peut être aussi simple qu’un changement de monomères d’ADN (c’est le cas dans cet exemple) ou aussi complexe qu’une réorganisation indépendante de systèmes multiples composés de nombreux gènes et membres du corps qui convergent sur une solution (c’est le cas du vol actif d’oiseaux et de chauves-souris). Dans les deux cas, cependant, nous pouvons déterminer qu’elles sont survenues par des événements indépendants sur des lignages séparés parce que ces caractéristiques ne correspondent pas à des événements uniques sur des arbres généalogiques des espèces.
Le pouvoir de la convergence
Puisque les homoplasies sont comme des marqueurs qui indiquent les événements évolutifs répétés, chercher des homoplasies dans les arbres généalogiques des espèces est utile pour tester des hypothèses sur la reproductibilité de l’évolution, ou sur la fréquence à laquelle des espèces convergent sur des solutions similaires. Il se trouve que l’évolution se répète beaucoup pour nombreux traits généraux. Il existe de nombreux exemples d’innovations répétées et indépendantes au cours de l’histoire évolutive, et nous en examinerons quelques uns dans les billets à venir :
- Forme aérodynamique du corps : les formes aérodynamiques du corps de ceux qui ont une vie aquatique comme les poissons, les ichtyosaures, les baleines, les phoques et les oiseaux plongeurs (les pingouins) sont toutes des adaptations indépendantes et convergentes à la vie aquatique.
- Vol actif : en plus des oiseaux et des chauves-souris, le vol actif a aussi évolué indépendamment chez les insectes et les ptérodactyles.
- Echolocalisation : certains mammifères comme les chauves-souris et les baleines ont indépendamment développé des systèmes qui leur permettent de localiser de la nourriture à travers la détection des échos du son qu’ils génèrent sur les structures et les proies de leur environnement.
- Yeux camérulaires : l’évolution répétée des yeux camérulaires (des yeux qui utilisent une lentille) est l’un des exemples les plus frappants de l’évolution convergente. Les yeux camérulaires ont évolué indépendamment chez les cnidaires (certaines méduses), les céphalopodes (comme les calmars et les pieuvres) et les vertébrés (oiseaux, mammifères).
Il faut noter que ces exemples répandus de convergence sont tous formés par l’environnement physique des organismes en question : la perception de la lumières (les yeux), la possibilité de voler dans les airs (les ailes), ou de se déplacer efficacement dans l’eau (corps aérodynamique). On pourrait s’attendre à ce que la présence fixe de ces caractéristiques environnementales conduise à l’adaptation de nombreuses espèces.
Plus profondément
Dans le prochain billet de cette série, nous examinerons de plus près certains exemples de l’évolution convergente au niveau moléculaire, et verrons que dans certains cas, il y a en fait une homologie qui se tapit loin dessous certains événements convergents.
48 Articles pour la série :
- 01-L'évolution expliquée : Introduction
- 02-L'Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l'homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive (Cet article)
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l'humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour




{kind=link}
{kind=link}