
Le génome comparé à un texte ancien (4)
Dans le billet précédent, nous avons vu qu’à une large échelle d’organisation, le génome humain a les caractères qu’il devrait avoir s’il partageait un ancêtre avec d’autres grands singes. En continuant avec notre analogie du « livre », nous comparons maintenant ces textes à un niveau un peu plus détaillé : celui des phrases et des mots.
Comparaison des génomes au niveau des “phrases”
Dans un billet précédent, nous avons comparé les séquences ADN d’un gène que l’on trouve dans plusieurs espèces de drosophiles. De telles comparaisons sont aussi possibles en utilisant les séquences ADN de mammifères (humains et autres primates), et le patron qu’ils produisent est désormais familier :

Comme nous l’avons vu dans les séquences des drosophiles, ce gène est quasiment identique dans un certain nombre d’espèces. Typiquement, la séquence humaine et les séquences de trois autres primates (chimpanzés, gorilles et orang-outans) ne diffèrent que par quelques nucléotides (tout au plus 4 sur les 90 sont différents). De plus, comme nous l’avons vu auparavant, aucun besoin biologique ne rend cette identité nécessaire – en fait, même dans la petite région de ce gène, il y a plus de 53 millions façons différentes de coder la même séquence d’acides aminés ! Et pour aller encore plus loin, ce gène n’a pas particulièrement besoin, biologiquement, d’avoir exactement la même séquence d’acides aminés que l’on voit partagée par les primates. Dans d’autres organismes (comme ceux des chiens et des loups), une séquence un peu différente accomplit la même tâche.
Bien entendu, il n’est pas possible de montrer des alignements d’ADN trop longs sous ce format. Ce petit segment de gène, cependant, est représentatif des gènes (et même de génomes entiers) des primates. Une comparaison détaillée de toutes les séquences de gènes entre les humains et les chimpanzés, par exemple, révèle qu’elles sont identiques à 99,4% sur 1.85×107 (18 millions) nucléotides. Notez que les régions du génome qui codent les gènes sont une toute petite minorité des séquences du génome – les humains et les chimpanzés ont plus de 3,0×109 (3 milliards) de nucléotides dans leur génome. Parmi ces 3 milliards de nucléotides, 2,7 milliards d’entre eux s’alignent les uns aux autres, avec une différence de seulement 1,23%. Bref, quand on compare les séquences d’ADN des humains et des autres primates, on voit exactement le patron que l’on aurait prédit si l’on suppose un ancêtre commun – un patron consistant, avec quelques modifications d’un génome ancestral.
A la recherche de fautes de frappe
Dans un billet précédent, nous avons vu comment la réplication de l’ADN est un processus très précis, mais non parfait. Ces deux caractéristiques de la réplication de l’ADN signifient que des mutations peuvent affecter des gènes lorsqu’ils sont copiés, et que des copies futures faites à partir d’un modèle muté transmettront fidèlement cette mutation (du moins jusqu’à une seconde mutation au même endroit). Cela signifie que les séquences du gène peuvent persister dans les génomes encore longtemps après leur mutation si cette mutation ne fait pas perdre la fonction en question. (Si une mutation est désavantageuse, alors la sélection naturelle aura tendance à l’éliminer de la population, comme nous l’avons déjà vu.)
Regardons l’exemple d’un gène qui code pour une enzyme (L-gulonolactone oxidase, ou “GULO”) intervenant dans la synthèse de la vitamine C chez les mammifères. La plupart des mammifères fabriquent leur propre vitamine C à partir d’autres éléments de leur alimentation, et le gène GULO est nécessaire dans la dernière étape du processus qui convertit un précurseur de vitamine C en produit final. Comme nous l’avons vu pour d’autres gènes, la séquence de ce gène est conservée entre les mammifères – il a une séquence quasiment identique, maintenue à travers la sélection naturelle. Par exemple, une part de ce gène chez la vache (Bos taurus), le chien (Canis lupus) et le rat (Rattus norvegicus) a la séquence suivante (avec des différences par rapport à la séquencede la vache, surlignées en noir) :

Dans ces trois espèces, ce gène est fonctionnel, et toutes trois peuvent fabriquer leur propre vitamine C sans l’obtenir directement de leur alimentation.
Les humains, bien entendu, ne peuvent pas fabriquer leur propre vitamine C – nous attrapons le scorbut si nous n’obtenons pas de vitamine C de notre alimentation. Cette situation atypique (pour les mammifères) est partagée par d’autres grands singes, et pour la même raison. Bien que ces espèces aient une partie de la séquence ADN du gène GULO, celle-ci contient de nombreuses mutations qui rendent le gène incapable de produire une enzyme fonctionnelle. La même région du gène GULO montrée dans la figure ci-dessus a les séquences suivantes chez les humains, les chimpanzés et les orang-outans (cette fois avec des différences par rapport à la séquence humaine, surlignées en noir) :
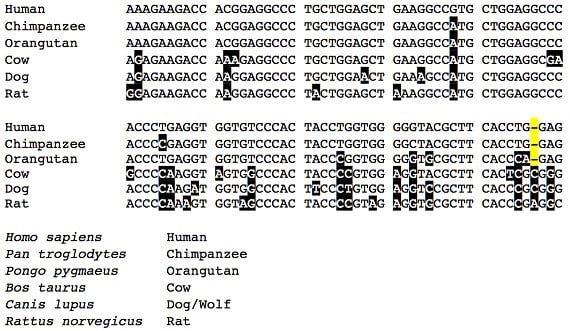
Une fois de plus on remarque que les séquences des primates sont quasiment identiques. Une nouvelle caractéristique à noter ici, cependant, c’est que ces trois copies du gène GULO ne sont pas fonctionnelles en partie parce qu’elles ont une délétion – un nucléotide en moins (surligné en jaune dans les séquences des primates). Cette délétion est identique chez les trois espèces, ajoutant une preuve à l’hypothèse de la « faute de frappe partagée » et copiée d’un premier texte – ou en termes biologiques, une délétion arrivée un jour chez l’ancêtre commun des humains, des chimpanzés et des orang-outans, ensuite héritée par les trois espèces. Les chiens, les vaches et les rats, cependant, ont bifurqué du lignage qui a conduit aux primates avant que cet événement de délétion ait eu lieu :
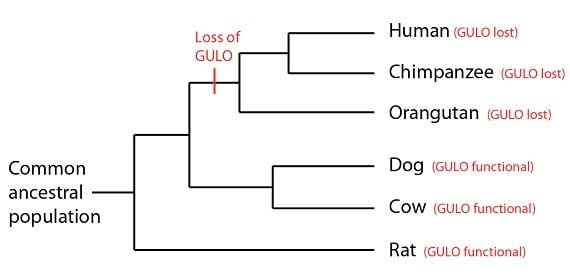
La perte de la fonction GULO ne semble pas avoir été un désavantage sélectif pour les primates à cette époque – probablement parce qu’ils avaient un régime alimentaire riche en vitamine C. En effet, même pour des humains, cette perte n’est pas un problème sérieux à moins que l’on ne se trouve sans source de vitamine C pour une période prolongée.
Le nez sait
Aussi intéressant que soit l’exemple du GULO (et c’est un exemple que j’ai déjà discuté plus en détail dans un autre contexte), ce n’est qu’un exemple de mutation partagée et identique parmi beaucoup d’autres que l’on trouve dans le génome humain et d’autres primates. Une étude qui a examiné en détail les mutations partagées par les primates s’est intéressée aux mutations dans les gènes consacrés au sens olfactif. Ces gènes codent pour des protéines que l’on trouve dans la membrane de cellules de l’épithélium nasal des mammifères, et que l’on appelle les récepteurs olfactifs. Les récepteurs olfactifs s’attachent aux éléments présents dans l’air, changent de forme lors du processus, et signalent ce changement au système nerveux produisant ainsi le sens olfactif.
Les nombreux récepteurs olfactifs qui agissent de concert sont ce qui donne à un parfum donné ses caractéristiques distinctives. Chez les mammifères, les gènes des récepteurs olfactifs constituent une part disproportionnée de leur génome, probablement parce que de tels gènes sont utiles pour trouver de la nourriture, un partenaire et en général pour percevoir son environnement. Malgré leur utilité, ces gènes peuvent aussi mutés et être perdus ; et en effet, le génome humain montre que notre espèce en a perdu plusieurs à cause de la mutation. Comme pour le gène GULO, cependant, ces séquences défectueuses de gène olfactif persistent sous une forme reconnaissable. Ce qui est plus important pour nous, cependant, c’est le patron que ces gènes mutés forment lorsqu’on les compare à des génomes d’autres primates. Comme nous l’avons introduit avec notre analogie du livre copié, nous nous attendons à trouver des fautes de frappe partagées entre les textes, et d’autres uniques à une édition. Pour les gènes olfactifs défectueux, nous observons précisément ces deux catégories : des mutations partagées, et des mutations uniques :
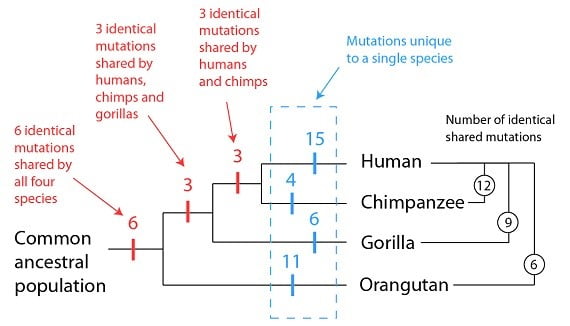
Comme vous pouvez le voir sur le diagramme ci-dessus, les humains partagent plus de gènes olfactifs identiques avec les chimpanzés, moins avec les gorilles et encore moins avec les orang-outans. Des 12 mutations qui sont identiques entre les humains et les chimpanzés, 9 sont aussi identiques avec les gorilles, et 6 avec les orang-outans. Ces mutations partagées et le patron que l’on trouve sont expliqués facilement par l’existence d’un ancêtre commun, comme indiqué en rouge dans le diagramme. Les mutations uniques à une espèce donnée sont aussi facilement expliquées comme survenant dans des populations séparées (en bleu).
Il est important de noter aussi ce que nous ne voyons pas lorsqu’on compare ces mutations entre les primates. Nous n’observons pas de mutations identiques entre les humains et les gorilles, par exemple, sans les trouver chez les chimpanzés. Cela est se comprend si la population ancestrale commune aux humains et aux gorilles est aussi celle aux humains, aux gorilles et aux chimpanzés. De même, si nous observons des mutations identiques partagées entre les humains et les orang-outans, nous pouvons prédire avec confiance que nous les observerons aussi chez les gorilles et les chimpanzés ; et de fait, nous les observons. Ce patron de mutations partagées est précisément celui que l’on prédirait s’il était produit par un ancêtre commun.
De multiples lignes d’évidence, une conclusion
Dans le prochain billet de cette série, nous reviendrons sur tout cela pour voir comment les multiples lignes d’évidence génomique pour l’évolution humaine forment un patron cohérent.
[p1]C’est normalement Canis lupus
48 Articles pour la série :
- 01-L'évolution expliquée : Introduction
- 02-L'Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l'homme
- 17-Le génome comparé à un texte ancien (4) (Cet article)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l'humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour




