
![Photo credit: Flickr user edenpictures [CC BY 2.0]](data:image/svg+xml;base64,PHN2ZyB4bWxucz0iaHR0cDovL3d3dy53My5vcmcvMjAwMC9zdmciIHdpZHRoPSI1NzAiIGhlaWdodD0iMzUxIiB2aWV3Qm94PSIwIDAgNTcwIDM1MSI+PHJlY3Qgd2lkdGg9IjEwMCUiIGhlaWdodD0iMTAwJSIgc3R5bGU9ImZpbGw6I2EzNzlhZjtmaWxsLW9wYWNpdHk6IDAuMTsiLz48L3N2Zz4=)
- Photo credit: Flickr user edenpictures [CC BY 2.0]
![Photo credit: Flickr user edenpictures [CC BY 2.0]](https://scienceetfoi.com/wp-content/uploads/2015/01/12.jpg)
La mise en place des plans d’organisation des vertébrés : cinquième partie.
Dans ce billet, nous verrons l’origine et la diversification des mammifères, en expliquant les éléments génomiques qui montrent la transition d’un œuf rempli de vitellus (c’est-à-dire le jaune d’œuf) à une alimentation de l’embryon par le placenta.
Comme nous l’avons vu dans le dernier billet de cette série, les tétrapodes du groupe couronne sont un groupe très divers, allant des amphibiens aux reptiles, aux oiseaux et aux mammifères. Puisque ce dernier groupe inclut notre propre espèce, il serait dommage de ne pas examiner au moins quelques caractères qui distinguent les mammifères des autres tétrapodes, ainsi que l’histoire de leur évolution.
Le groupe souche des mammifères : les synapsides.
Les mammifères sont les seuls représentants en vie du groupe des synapsides, groupe qui s’est séparé du lignage des dinosaures/aviens (les sauropsides) et qui a continué à se diversifier à partir de la période carbonifère tardive, il y a environ 325 millions d’années. Les nombreux synapsides que l’on connaît par les données fossiles sont des groupes souche des mammifères, c’est-à-dire des organismes apparentés aux mammifères qui se sont séparés du lignage qui a conduit au groupe couronne des mammifères (la dernière population ancestrale commune de tous les mammifères en vie, et toutes les espèces descendant de cette population). Le fait que certains de ces groupes souche, les synapsides éteints, aient été appelés « reptiles type mammifère » et d’autres « mammifères type reptile » révèle la nature transitionnelle de leurs caractères, qui brouillent la distinction entre les reptiles et les mammifères tels qu’ils nous sont connus aujourd’hui : c’est-à-dire à travers l’accumulation graduelle de traits caractéristiques du groupe couronne des mammifères, et par un patron qui indique dans quel ordre ces caractéristiques ont été acquises. On peut prendre pour exemples la morphologie de la mâchoire (qui inclut l’exaptation des os de la mâchoire en une fonction spécialisée, l’ouïe, dans l’oreille interne), le développement des cheveux, et la lactation (la sécrétion de lait pour nourrir les petits).
De l’oeuf au placenta
Il m’arrive parfois de rencontrer des non biologistes (et même certains biologistes) qui sont étonnés d’apprendre que la viviparité n’est pas une caractéristique qui définit les mammifères (ou plus précisément, les mammifères du groupe couronne). La raison en est qu’un lignage de mammifères qui pondent des œufs, les monotrèmes, a des représentants en vie. Le fait que des mammifères qui pondent des œufs existent aujourd’hui signifie qu’ils font partie du groupe couronne (par définition), et en tant que tels, toute caractéristique qu’ils n’ont pas ne peut pas définir le groupe couronne :

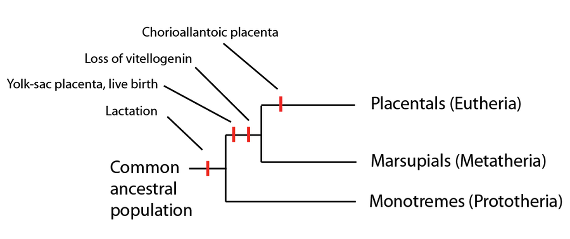
Les mammifères du groupe couronne incluent des mammifères qui pondent des oeufs (les monotrèmes) comme des mammifères qui n’en pondent pas (les marsupiaux et les mammifères placentaires). Parmi les caractères signalés sur cette phylogénie, seule la lactation est une caractéristique commune au groupe couronne tout entier.
Chez les mammifères ovipares, comme l’ornithorynque et les différentes espèces des échidnés, une fois l’éclosion passée, les petits sont nourris par le lait de leur mère, sécrété à certains endroits de sa peau qu’ils lèchent. Chez les marsupiaux, la période de gestation est bien plus courte que chez les mammifères placentaires, et après leur naissance les petits (dans un état encore très proche de l’embryon) rampent jusqu’à une poche protégée où ils se nourrissent à une mamelle pour compléter leur développement. Chez les marsupiaux, une brève connexion se forme in utero entre l’embryon et la mère à travers une membrane dite sac vitellin pour former un omphalo-placenta. Eh oui, il est étonnant que les marsupiaux et les mammifères placentaires, c’est-à-dire les euthériens, aient tous les deux un placenta. La différence est que les mammifères placentaires forment leur placenta à partir d’une membrane différente, la membrane allantoïde (allantoplancenta).
Si les monotrèmes et les marsupiaux ne sont pas des groupes souche des mammifères (puisque leur lignage persiste jusqu’à aujourd’hui), nous pouvons apprécier leurs caractères de la même manière que nous l’avons fait pour les groupes souche. (Autrement dit, si les lignages des monotrèmes et des marsupiaux s’étaient éteints, nous appellerions les euthériens « mammifères » et les monotrèmes et les marsupiaux seraient des groupes souche sur le lignage euthérien.) Comme nous l’avons vu avec d’autres groupes souche, les monotrèmes et les marsupiaux démontrent que l’état euthérien a été atteint pas à pas, au cours du temps. Bien que ce processus ait été en réalité graduel, nous pouvons arbitrairement dénoter certaines « étapes » :
- Ponte des oeufs suivie d’une lactation post-éclosion (état monotrème)
- Courte gestation suivie d’un sac vitellin et d’une lactation post-natale (état des marsupiaux)
- Gestation avec une combinaison de l’omphalo-placenta et de l’allantoplacenta (avec extension du temps de gestation) suivie d’une lactation post-natale
- Réduction de l’omphalo-placenta en faveur de l’allantoplacenta (avec une extension plus grande du temps de gestation) suivie d’une lactation post-natale
- Longue gestation avec un allantoplacenta suivie d’une lactation post-natale (condition euthérienne).
Au fil du temps, les métathériens et les euthériens se seraient éloignés de la nutrition par la vésicule vitelline pour leurs embryons in utero, et se seraient dirigés vers une nutrition faisant intervenir le placenta. De cette manière, on peut prédire que la mécanique biochimique qui permet la nutrition par la vésicule vitelline serait devenue de moins en moins importante, et finalement, inutile. Au niveau génétique, les gènes requis pour la production du sac vitellin n’auraient plus contribué à terme à la survie ou à la reproduction de l’organisme, ce qui signifie qu’ils n’auraient plus été sélectionnés, auraient été désormais libres d’accumuler des mutations sans conséquences pour l’organisme.
A la recherche du mort parmi les vivants
En termes pratiques, lorsqu’un gène n’est plus sous sélection naturelle, il est alors maintenu par la précision globale de la réplication de l’ADN lors de la division des cellules. Si la réplication ADN est plutôt exacte, elle n’est pas parfaite. Pour des séquences sous sélection naturelle, les mutations sont éliminées de la population si les individus qui portent ces variantes ne peuvent pas se reproduire à la même fréquence que leurs parents qui ne portent pas cette mutation. En ce qui concerne les gènes qui ne sont plus sous sélection naturelle, les mutations vont s’accumuler lentement au cours du temps.
En ce qui concerne les marsupiaux et les mammifères placentaires, on s’attendrait à ce qu’un tel gène, la vitellogénine, soit abandonné par la sélection une fois le placenta établi. La vitellogénine est vitale pour la formation du vitellus (ou jaune d’œuf), puisqu’il est un porteur majeur de nutriments qui partent du foie jusqu’au vitellus en formation chez les organismes ovipares. En 2008, un groupe de recherche a cherché les résidus des séquences de la vitellogénine chez les mammifères placentaires, les marsupiaux, et les monotrèmes. Les monotrèmes, comme vous vous y attendez, ont des séquences génétiques fonctionnelles de la vitellogénine, puisqu’ils sont des mammifères ovipares. Si les marsupiaux et les mammifères placentaires n’ont pas de séquences génétiques fonctionnelles de la vitellogénine, ils possèdent cependant les résidus des séquences mutées de la vitellogénine, indiquant que ces lignages ont autrefois possédé une mécanique biochimique fonctionnelle pour transférer des nutriments au vitellus. Cette observation correspond parfaitement à la prédiction phylogénétique selon laquelle les mammifères placentaires et les marsupiaux partagent une population commune avec les monotrèmes (et bien sûr d’autres tétrapodes) – l’état ancestral étant ovipare, caractéristique par la suite perdue dans la population commune des marsupiaux et des mammifères placentaires :
Le séquençage du génome nous permet donc de tester des prédictions spécifiques à propos de ce que nous devrions découvrir (basées sur des phylogénies assemblées qui utilisent des caractères anatomiques et morphologiques). Dans ce cas, la présence de séquences de vitellogénine chez les mammifères placentaires (génome humain compris) qui ne peuvent pas fonctionner pour faire le vitellus est un exemple frappant d’une prédiction confirmée sur l’évolution (prédiction qui continue à être très problématique pour les groupes anti-évolution). Dans ce contexte, cependant, la perte de la vitellogénine n’était qu’une petite étape sur le chemin vers les lignages métathérien et euthérien, qui, au contraire des monotrèmes, demeurent prospères aujourd’hui.
Dans le prochain billet de cette série, nous explorerons la diversification des mammifères placentaires, en incluant le lignage qui conduit à notre propre espèce : les primates.
48 Articles pour la série :
- 01-L'évolution expliquée : Introduction
- 02-L'Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l'homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères. (Cet article)
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l'humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour



