
Dans le billet précédent, nous avons introduit l’idée que les espèces peuvent se former dans le même lieu géographique par le partage des ressources – lorsque les populations exploitent avec le temps différentes niches,. Dans ce billet, nous explorerons ce phénomène en détail, en utilisant l’exemple d’une espèce émergente qui s’est formée dans un passé très récent, et sous l’observation de l’homme : la diversification des mouches de la pomme, Rhagoletis pomonella. Ces mouches sont attirées par les fruits de l’aubépine quand ils ne sont pas encore mûrs, parent sauvage des pommiers domestiques. Le fruit de l’aubépine est aussi le lieu où les mouches de la pomme trouvent leur partenaire et pondent leurs œufs, afin de permettre aux larves de se nourrir du fruit (ce qui le gâte et le fait tomber plus tôt, en emportant la larve). Les mouches de la pomme ne produisent qu’une génération par an, et survivent à l’hiver sous forme de chrysalides. De plus, elles ont une durée de vie d’adulte courte, ce qui ne leur donne qu’une petite période pour trouver un partenaire, se reproduire, et pour pondre les œufs en ce qui concerne les femelles. Cette période cruciale correspond bien sûr au cycle de vie de l’aubépine – lorsque ses fruits peuvent être une source alimentaire et un lieu de rencontre. En tant que telle, la sélection naturelle (exercée par le cycle de vie de l’aubépine) agit sur la variation génétique qui contrôle le temps d’éclosion chez les populations de mouches de la pomme. Le moment de l’éclosion est sous contrôle héréditaire et les mouches qui éclosent alors que la saison des fruits de l’aubépine touche à sa fin (ou pire, quand elle est finie et qu’il n’y a plus du tout de fruits) ne se reproduisent pas avec autant de succès que les mouches qui éclosent lorsqu’il y a abondance de fruits. Sans surprise, nous observons des populations de mouches de la pomme bien synchronisées avec les plantes qui les accueillent : la plupart des membres de n’importe quelle population de mouches éclot de concert avec la quantité de fruits disponibles :
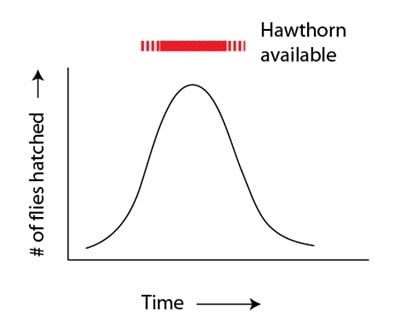
Le temps d’éclosion est un exemple de caractère continu, par opposition au caractère discret. Le caractère discret possède des catégories distinctes : les yeux bleus et les yeux noirs, ou les fleurs rouges et les fleurs blanches, etc. Nombre de ces caractères ne peuvent pas entrer dans de telles catégories et sont plutôt distribués dans les populations. La taille et le poids sont des exemples de caractères continus, ainsi que le temps d’éclosion des mouches de la pomme. L’effet que l’arbre d’aubépine a sur la mouche de pomme est un exemple de la sélection stabilisante – les mouches qui éclosent en début ou en fin de saison, alors qu’il n’y a pas encore beaucoup de fruits, ne sont pas sélectionnées. Cette sélection a pour effet global de synchroniser l’éclosion des mouches avec la disponibilité du fruit, génération après génération.
Tentée par une pomme
Quelque chose est arrivé et a perturbé cette interaction stable et équilibrée : l’introduction de du pommier domestique en Amérique du Nord par les colons européens. Comme nous l’avons noté ci-dessus, l’aubépine et le pommier appartiennent à la même famille de végétaux, et leurs fruits sont similaires. Mais le moment auquel le fruit se développe est différent : les pommiers domestiques produisent des fruits quelques semaines plus tôt que les aubépines. L’introduction de pommiers dans l’habitat des mouches de pomme a ainsi donné une source alimentaire potentielle aux mouches qui éclosent trop tôt :
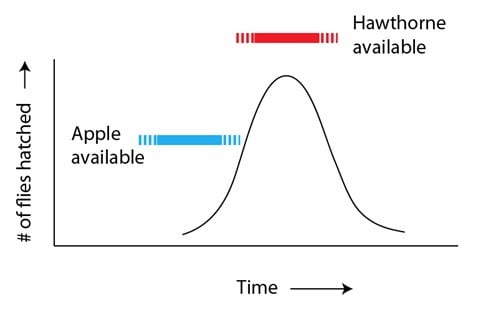
Le résultat pour ces mouches “précoces”, attirées par ce fruit nouveau dans leur environnement, mais similaire à celui qu’elles connaissent, revêt deux aspects : (a) il permet d’avoir accès à une source alimentaire avec une moindre compétition de la part des membres de leur propre espèce et (b) il s’agit de trouver un partenaire aux tendances similaires, c’est-à-dire attiré par la pomme. Ce qui était au départ une combinaison génétique « perdante » (éclosion trop précoce, sans suffisamment de nourriture ou de perspective de trouver un partenaire) devient une combinaison « gagnante ». Il en résulte que la variante « précoce » peut désormais se reproduire plus effectivement qu’auparavant, et ainsi augmenter en nombre à travers les générations successives :
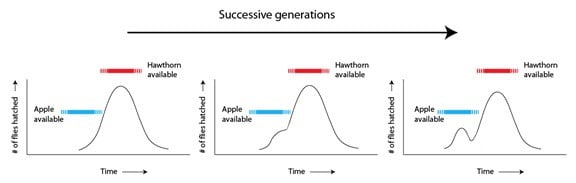
Autrement dit, une fois que les pommes sont présentes, l’environnement ne sélectionne plus les populations de mouches de façon stabilisante, mais fait plutôt d’une variation deux sous-populations. La sélection devient diversifiante. Ce qu’il est important de noter, c’est que ces deux sous-populations ne se diversifient pas seulement par rapport au moment d’éclosion et à la préférence du fruit, mais aussi (étant donnée la nature de leur biologie) par rapport à leur préférence en matière de partenaire. Alors que les variantes « pommes » augmentent en nombre, les mouches se reproduisent naturellement plus fréquemment avec d’autres variantes « pommes », puisqu’elles rencontrent leur partenaire sur des pommiers. Il en résulte une barrière partielle au flux d’allèles qui renforce les différences émergentes entre les deux groupes à travers le temps.
Si les espèces de Rhagoletis pomonella de l’aubépine et de la pomme ont intéressé les hommes depuis des siècles (surtout à cause de l’impact économique de la mouche de la pomme en tant que parasite), les généticiens commencent juste à comprendre les différents allèle qui ont été les cibles de la sélection lors du processus de séparation. Des gènes que l’on connaissait par des recherches antécédentes comme affectant le temps d’éclosion s’avèrent avoir des allèles différents dans les deux groupes. D’autres gènes candidats incluent le récepteur de protéines que les mouches utilisent pour détecter les odeurs de leur cible, les fruits – certains allèles sont plus ajustés aux odeurs de la pomme, d’autres aux odeurs de l’aubépine. Ce qui a commencé comme étant une variation à l’intérieur d’une population est désormais partagé par la sélection en combinaisons d’allèle adaptées à des niches distinctes – et étant donné la rapidité avec laquelle s’est faite le changement pour la pomme, il est probable que de nouvelles mutations n’aient pas joué de rôle. La recombinaison et la ségrégation d’allèles existant déjà sur de nombreux gènes ont été suffisantes pour donner des différences génétiques qui correspondaient à certains membres de la population d’origine et qui permettaient d’exploiter une nouvelle opportunité. Il en a résulté le changement de certains caractères continus (le temps d’éclosion, la préférence pour une odeur de fruit) correspondant à une nouvelle niche environnementale et ériger une barrière au flux d’allèles.
Une fois examinés les gènes (et leurs allèles) sélectionnés pendant cet événement de spéciation, il y a plusieurs choses à noter. Le nombre de gènes sous sélection (et ainsi possédant différents allèles dans les deux nouvelles espèces) est relativement faible. Seuls les allèles qui affectent les caractères pertinents à l’adaptation à la nouvelle niche sont sélectionnés. La plupart des gènes demeurent identiques entre les deux populations, puisqu’elles ne sont pas passées par une sélection diversifiante mais par une sélection stabilisante pour leurs rôles (identiques) dans les deux espèces. Par exemple, considérons les gènes requis pour la conversion de l’énergie, ou pour le développement d’une aile, processus dont les deux espèces ont toujours besoin de la même façon. Ces gènes ont les mêmes allèles (ou peut-être un seul allèle) dans les deux populations, puisque la fonction de ces gènes n’était pas pertinente pour l’adaptation à la nouvelle niche. En quelques mots, le schéma général que produit la spéciation consiste en un petit nombre de différences d’allèles pour les gènes sous sélection (ou les gènes qui ont expérimenté la dérive pas hasard) avec la grande majorité de gènes identiques qui n’ont pas été soumis à la sélection ou à la dérive).
En effet, l’une des raisons pour lesquelles nous pouvons croire que les “spécialistes” des pommes et des aubépines de Rhagoletis pomonella sont en fait les produits d’un événement de spéciation récent (mis à part le fait que des fermiers ont observé l’émergence des deux espèces), c’est la ressemblance qui existe entre leur génome – ils n’ont que de petites différences dans un petit nombre de gènes. Biologiquement, savoir si elles sont en fait deux espèces séparées demeure une question ouverte, puisqu’elles continuent à échanger des allèles, bien qu’à un taux beaucoup plus faible qu’à l’intérieur de leurs populations respectives. Comme nous l’avons vu dans le cas des espèces en anneau, cet exemple nous montre qu’il est possible d’observer aujourd’hui encore les caractéristiques que nous pourrions prédire dans le cas d’un événement de spéciation en cours. De plus, il montre qu’un petit nombre de différences, dérivées de variations qui existaient déjà dans la population, peut mettre deux sous-populations sur une trajectoire qui augmente progressivement la barrière de flux d’allèles entre elles. Avec le temps, ces effets peuvent amener à la formation d’espèces proches.
A long terme
La production d’espèces proches à partir d’une population ancestrale commune est un sujet peu controversée chez les chrétiens évangéliques, bien que les mécanismes sous-tendant de tels événements ne sont souvent pas appréciés. Ce qui est plus controversé pour beaucoup, cependant, est la suggestion que ces mécanismes produisent aussi des espèces largement divergentes dans un temps long. Dans le prochain billet, nous examinerons certains arguments qui soutiennent l’hypothèse selon laquelle des espèces modernes très diverses sont effectivement dérivées d’une population ancestrale, dans un passé très lointain.
Pour en savoir plus
Schwarz, D. et al., (2009). Sympatric ecological speciation meets pyrosequencing: sampling the transcriptome of the apple maggot Rhagoletis pomonella. BMC Genomics 10; 633. (http://www.biomedcentral.com/1471-2164/10/633)
48 Articles pour la série :
- 01-L'évolution expliquée : Introduction
- 02-L'Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4) (Cet article)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l'homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l'humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour




