Dans le dernier billet de cette série, nous avons examiné le cas relativement simple de la séparation géographique des populations (par exemple lorsqu’une population est fondée sur une île). La séparation géographique est une barrière effective à ce que les biologistes appellent « le flux des gènes » entre les populations ; un effet qu’on pourrait décrire de manière plus adéquate par l’expression « flux d’allèles ». Alors que de nouveaux allèles surviennent dans des populations séparées, l’absence de reproduction entre les populations a pour conséquence de garder chaque allèle dans la population à l’intérieur de laquelle il est survenu. Ces nouveaux allèles peuvent contribuer à la spéciation à travers le temps, s’ils affectent les caractéristiques de l’organisme. Si, à l’autre extrême, de nouveaux allèles peuvent passer librement entre les deux populations, alors ils ne contribueront pas à un événement de spéciation, puisqu’ils ne rendront pas les populations différentes dans le temps.
D’une espèce à l’autre
Si ces deux extrêmes (les populations géographiquement séparées et les populations continues) sont aisées à comprendre, il est possible de trouver des situations intermédiaires. Par exemple, considérons deux populations (que nous appellerons A et B) qui sont membres de la même espèce. Elles peuvent échanger des allèles, mais à un rythme réduit par comparaison au partage qui se fait à l’intérieur d’une population. Cet effet peut survenir à cause de la forme géographique de leur habitat : s’il est long et étroit, alors les deux populations peuvent n’être contiguës que sur une petite partie de leur étendue. Cela signifie qu’en moyenne, un individu de la population A trouvera plus probablement un partenaire dans la population A que dans la population B, dans la petite région qu’ils partagent. Nous pouvons représenter ceci par des boîtes qui représentent les deux populations, contiguës l’une à l’autre par un seul côté, étroit :
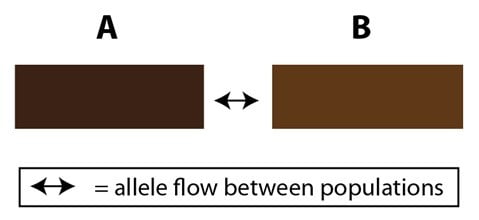
Cet arrangement restreint ainsi, sans l’abolir complètement, le flux d’allèles entre les deux populations. Il constitue une barrière partielle au flux d’allèles. Les populations A et B sont les membres d’une même espèce mais les deux populations ne sont pas génétiquement identiques. Alors que de nouveaux allèles surviennent dans la population A, ils ne sont pas partagés avec la population B aussi souvent qu’à l’intérieur de la population A, et vice versa. Il est aussi possible que les deux populations expérimentent des différences dans la sélection naturelle (puisque leurs environnements ne sont pas identiques), et/ou des différences dans la dérive génétique, en fonction de la taille de la population. Le résultat net est un équilibre des forces qui agissent sur les deux populations ; certaines favorisant les différences (la sélection et/ou la dérive) et une autre favorisant les similitudes (un flux limité d’allèles par la reproduction entre les populations).
Dans la nature, cet effet peut s’étendre à des populations multiples sur un « fil » qui s’étend sur une suite d’habitats adaptés. Ajoutons trois populations (C, D et E) à l’exemple précédent pour l’illustrer :
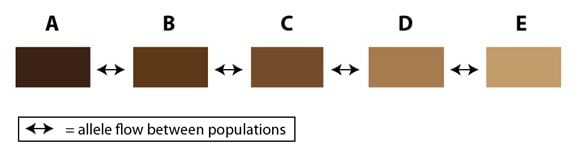
Une fois que les populations s’étendent sur une large région géographique, les différences entre les populations aux extrémités (les populations A et E dans notre diagramme) peuvent devenir significatives. Dans certains cas, et c’est intéressant, les populations des extrémités peuvent être assez différentes de sorte qu’elles ne se reconnaissent plus comme membres de la même espèce, malgré leur connexion génétique par une série de populations intermédiaires. Dans certains cas, les scientifiques ont besoin de rassembler des membres des populations aux extrémités pour voir s’ils peuvent se reproduire entre eux (c’est-à-dire en employant le concept biologique d’espèce). Dans d’autres cas, la topographie de l’habitat les rassemble dans la nature, ce qui permet aux populations aux extrémités du fil de se rencontrer en formant un anneau, mais avec une barrière naturelle au milieu (comme une montagne ou une vallée dont l’habitat est inadapté). Le résultat est ce que l’on appelle la « variation clinale » (ring species = espèce en anneau) :
On voit la difficulté inhérente à la définition des espèces séparées (s’il y en a). Il y a un flux d’allèles entre toutes ces populations, mais seulement en anneau. Les deux populations des extrémités (qui se recouvrent), malgré leur rencontre dans le même habitat, sont assez différentes pour ne pas se reproduire entre elles. Définir ces populations comme des espèces différentes (ou pas), c’est vainement essayer de dessiner une frontière de démarcation dans un dégradé de couleurs. Pour ceux qui sont intéressés par un exemple réel d’une variation clinale, les sous-espèces de la salamandre Ensatina eschscholtzii sur la côte ouest de l’Amérique du nord en sont des cas classiques et demeurent encore actuellement un sujet de recherche.
Si nous rencontrions les populations des extrémités sans les intermédiaires, nous n’hésiterions pas à les classifier comme des espèces distinctes. Il est également aisé de voir ce qui suivrait si n’importe quelle population intermédiaire était perdue, ou si des changements dans l’habitat empêchaient la connexion entre n’importe quelles populations ; le résultat serait une interruption dans la chaîne de flux d’allèles, coupant les populations terminales les unes des autres. Ce que la variation clinale illustre, c’est que bien que la spéciation soit un lent processus d’accumulation de différences entre les populations, elle est possible même sans une barrière totale au flux des allèles.
La spéciation sans la séparation géographique
Si la variation clinale illustre la façon dont les espèces peuvent se former sur une large région géographique, des barrières au flux d’allèles peuvent aussi survenir à l’intérieur d’une population dans une localité plus compacte. Il suffit d’avoir un biais qui promeut l’échange des allèles à l’intérieur d’un sous-groupe de la population, au prix d’un moindre échange dans la population plus large ; et comme nous l’avons vu avec la variation clinale, cette barrière ne doit pas être absolue pour autoriser deux sous-populations à accumuler des différences entre elles, pour diverger ensuite à travers le temps. Par exemple, une sous-population peut commencer à exploiter différemment des ressources à l’intérieur d’une même aire géographique ; un effet que l’on appelle la partition ou la différenciation de niches. Alors que des sous-populations commencent à se spécialiser dans des « façons de vivre » à peine différentes, selon l’expression de Darwin, il devient plus probable qu’elles se reproduisent à l’intérieur de la sous-population plutôt que dans la population entière. Puisque la reproduction préférentielle est une barrière (partielle) au flux d’allèles, cela peut placer les sous-populations sur une trajectoire génétique qui renforce leurs différences et les amène à un événement de spéciation.
La partition de niches est le mécanisme probable conduisant à des événements de spéciation rapides et multiples qui arrivent lorsqu’une population fondatrice atteint un nouvel habitat, dans lequel les prédateurs sont largement absents. La colonisation d’îles volcaniques, un sujet que nous avons déjà discuté, peut amener à une radiation adaptative. Un exemple est celui des nombreuses espèces de pinsons dans les îles Galapagos qui viennent d’une seule espèce de pinsons, celle qui a colonisé l’archipel à l’origine. Par la suite elle s’est diversifiée en de nombreuses espèces qui se sont spécialisées dans des ressources alimentaires différentes. En l’absence d’autres oiseaux sur l’île, de nombreuses « façons de vivre » (que l’on appelle aujourd’hui des niches) étaient disponibles pour différentes sous-populations d’oiseaux.
En résumé : la spéciation commence par des barrières au flux d’allèles
La séparation géographique complète, la séparation géographique partielle vue avec la variation clinale, et le partage des ressources de sous-populations sont toutes des barrières au flux d’allèles entre les membres d’une même population (du moins au départ). Ceci permet à de nouveaux allèles de survenir, sans être partagés par les deux populations, et disjoint ainsi les caractéristiques moyennes des deux groupes. Dans le prochain billet, nous examinerons certains traits auxquels ces allèles contribuent ; des traits qui améliorent les barrières au flux d’allèles et permettent ainsi les événements de spéciation.
48 Articles pour la série :
- 01-L'évolution expliquée : Introduction
- 02-L'Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3 (Cet article)
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l'homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l'humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour