
Dans cette nouvelle série de billets, Dennis Venema explique les mécanismes qui sont en jeu dans la formation d’une nouvelle espèce, ce que l’on appelle la spéciation. On est ici au cœur de la théorie de l’évolution biologique, l’origine des espèces pour faire écho au le livre de Charles Darwin. Et comme nous le verrons tout se joue au niveau des populations. Bonne lecture !
– Pascal Touzet
De la variation à la spéciation (1)
Dans le dernier billet de cette série, nous avons examiné la façon dont la variation de l’ADN survient par des événements aléatoires, comme lorsque des paires sont inappropriées, ou lors de duplications ou de délétions. Afin de comprendre la façon dont cette variation peut contribuer à un événement de spéciation, nous verrons la manière dont la variation s’étend dans une population. D’abord, nous avons besoin d’un peu de vocabulaire pour faciliter la discussion ; plus spécifiquement, nous avons besoin d’expliquer la distinction entre le gène et l’allèle.
En tant que généticien, il m’arrive de m’arracher les cheveux lorsque je lis dans des journaux grand public des articles à contenu scientifique. L’une des choses qui m’agacent le plus par exemple, c’est l’utilisation du terme « gène » lorsqu’on dit qu’un individu « a le gène » d’un trait spécifique. Nous avons déjà dit que les gènes sont une section de séquence d’ADN sur un chromosome qui contribue à quelque fonction, habituellement en codant pour une protéine. Ce que nous n’avons pas, cependant, ce sont des gènes identiques – il y a des différences qui surviennent dans les erreurs de copie que nous avons déjà exposées. Ces différences sont appelées des allèles. Un allèle est comme une « version » ou une « saveur » d’un gène. Les événements mutationnels ne créent pas habituellement de nouveaux gènes (bien qu’ils le puissent par la duplication). Habituellement, ce sont de nouveaux allèles qui sont créés. Dans un billet précédent, nous avons utilisé les Lego d’enfants pour illustrer la façon dont une nouvelle variante peut survenir quand deux nucléotides forment par erreur une paire lors de la réplication du chromosome :
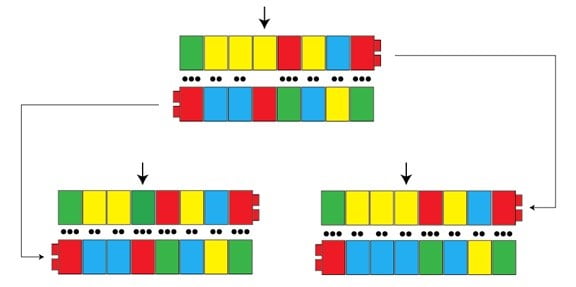
Dans cet exemple, nous avons une séquence qui, par une erreur de copie, devient deux différentes versions de ce qui est (presque) la même séquence. Ces différences sont appelées deux allèles distincts, et s’ils affectent la fonction d’un gène, ils peuvent avoir un effet significatif à l’échelle de tout l’organisme. Lorsque les journalistes parlent du « gène » pour tel ou tel trait, ce qu’ils veulent vraiment dire, c’est l’allèle pour tel ou tel trait, c’est-à-dire la variante spécifique d’un gène responsable d’une condition médicale spécifique, par exemple.
Sélection et dérive
La variation de l’ADN n’est rien d’autre que la production de nouveaux allèles – mais qu’arrive-t-il à ces allèles à travers le temps ? De toute évidence, lorsqu’un nouvel allèle survient, il n’est présent que dans un individu. S’il doit avoir un impact sur la population dans son ensemble, il faut qu’il s’étende à d’autres individus en se transmettant aux descendants. En ce sens, la variation peut entrer dans une population puis devenir plus commune. Il existe beaucoup de facteurs qui peuvent influencer ce processus. Si la taille de la population est petite, alors seule la chance peut augmenter (ou diminuer) la fréquence d’un allèle dans une population ; un effet que l’on nomme la « dérive génétique ». Puisque la dérive peut jouer un rôle majeur dans la façon dont les fréquences d’un allèle changent dans une population, il vaut la peine de prendre le temps de l’examiner en détail.
La dérive est essentiellement un événement d’échantillonnage non représentatif. Considérez une petite population d’organismes qui se reproduisent sexuellement, représentés par les rectangles, chacun contenant deux allèles (les carrés, les différences entre les allèles sont représentées par les couleurs). Chaque « reproduction » a une chance sur deux de transmettre l’un ou l’autre allèle à la prochaine génération. (Note : chaque événement de « transmission » est indépendant ; par exemple, il n’y a pas de mécanisme qui garantit qu’un individu transmette ses deux allèles s’il se reproduit deux fois.) En ce qui concerne la paire de gauche, un parent a deux allèles jaunes et l’autre un allèle bleu et un allèle jaune. Lorsqu’ils se reproduisent, par chance, le parent possédant les deux allèles ne transmet que le jaune aux deux descendants. Pour ce qui est de la paire de droite, le parent possédant les deux allèles transmet aussi l’allèle jaune les deux fois, et son allèle bleu pas du tout. Ces événements aléatoires font dériver significativement la fréquence des deux allèles au d’une génération à l’autre :
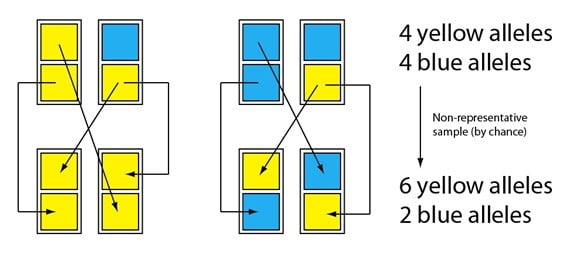
Imaginez maintenant que les descendants se reproduisent aussi et que cette fois encore, nous avons par chance un échantillon légèrement non-représentatif pour former la prochaine génération :
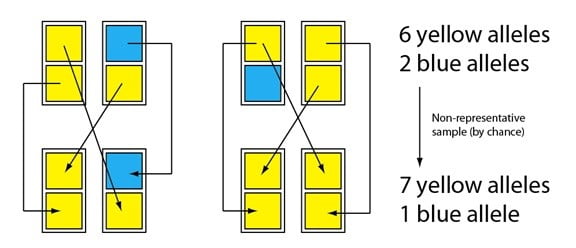
L’idée est que cette petite population est sujette à de larges fluctuations dans les fréquences des allèles bleus ou jaunes parce qu’elle est petite. Cette petite taille signifie que les événements aléatoires au sein d’une paire se reproduisant ont un large impact sur la population dans son ensemble. Lors de la prochaine génération, par exemple, l’allèle bleu pourrait être complètement perdu ; et une fois qu’il a disparu, il sera absent jusqu’à ce qu’il survienne à nouveau par un nouvel événement mutationnel, ou jusqu’à ce qu’il s’introduise dans la population par un individu immigrant d’une population dans laquelle il est toujours présent.
La situation est assez différente pour des populations plus grandes. Imaginez une population de 1000 individus, avec un total de 500 allèles jaunes et de 500 allèles bleus, distribués au hasard dans les individus. Lorsque cette population se reproduit, le rapport d’une chance sur deux ne variera jamais beaucoup d’une génération à l’autre. Dans cette population nombreuse, les événements aléatoires à l’intérieur d’une paire reproductrice sont une petite proportion de la population dans son ensemble, et en moyenne, la population reflètera la probabilité d’une chance sur deux pour n’importe quel allèle transmis.
Alors comment les fréquences de l’allèle peuvent-elles changer dans des populations de grande taille, alors que la dérve génétique est impuissante ? Nous avons déjà examiné un mécanisme qui peut l’accomplir : la sélection naturelle. La sélection naturelle est simplement l’effet de la reproduction plus fréquente des individus qui possèdent un certain allèle que de ceux qui ne le possèdent pas. Avec le temps, les allèles sélectionnés vont donc augmenter en fréquence dans la population. Par exemple pour les chiens, au début du processus de domestication, la duplication des gènes d’amylase est arrivée par un seul événement mutationnel aléatoire. Les chiens qui portaient l’allèle dupliqué de l’amylase se reproduisaient à une fréquence un peu plus élevée que ceux qui ne le portaient pas, puisque la duplication de l’allèle permettait aux chiens de consommer la nourriture qu’ils recevaient de leur nouvel environnement (c’est-à-dire des humains). Avec le temps, l’allèle dupliqué est devenu si fréquent dans la population canine que l’allèle ancestral et non dupliqué a été perdu. Le nouvel allèle était « fixé » dans la population : il avait une fréquence de 100%.
Pour résumer, dans une petite population, la dérive peut avoir un large impact sur les fréquences des allèles d’une génération à l’autre. Dans des populations plus nombreuses, la sélection naturelle prédomine, et la dérive a peu d’impact. Ces deux mécanismes peuvent contribuer au changement des fréquences d’allèles à travers le temps à l’intérieur d’une population, et en tant que tels, les deux peuvent être des facteurs qui contribuent à des événements de spéciation.
Changer la fréquence des allèles et la spéciation
La spéciation est la production de deux espèces à partir d’une population ancestrale. (Nous avons déjà dit que la définition de « l’espèce » est confuse ; le fait qu’elles surviennent lentement et progressivement les rend difficiles à définir.)
Une façon de comprendre la façon dont la spéciation commence, c’est de considérer deux populations de la même espèce qui pour quelque raison que ce soit, cessent de se reproduire entre elles ; peut-être à cause d’un isolement géographique. Si une « barrière » géographique n’a rien à voir avec les différences génétiques ou la compatibilité reproductive, si une telle barrière est en place, alors les allèles qui surviennent dans une population ne seront pas transférés dans l’autre population. De plus, si deux populations n’échangent pas d’allèles, alors les fréquences d’allèles dans les deux populations ne sont plus attachées l’une à l’autre et n’ont plus la même moyenne. Cela signifie que la dérive et la sélection naturelle agiront indépendamment dans les deux populations. Une fois que les populations sont séparées et isolées, elles peuvent suivre des trajectoires différentes – une population peut être d’abord petite, et être dominée par la dérive jusqu’à ce qu’elle augmente de taille. L’autre population peut rester nombreuse, et être sujette à la sélection naturelle d’une façon différente. Après une longue période, les deux populations peuvent devenir suffisamment différentes génétiquement pour former deux espèces. La clé, bien sûr, c’est la nature de la barrière qui empêche l’échange des allèles entre des populations. Dans le prochain billet de cette série, nous examinerons comment de telles barrières peuvent se former entre les populations.
48 Articles pour la série :
- 01-L'évolution expliquée : Introduction
- 02-L'Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1) (Cet article)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l'homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l'humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour



