
Article 29 sur un total de 48 pour la série :

Dans le dernier billet de cette série, nous avons introduit les caractères qui définissaient les chordés :
- une corde dorsale nerveuse vide
- une lamelle flexible que l’on appelle notochorde
- un pharynx (avec des sillons pharyngés, ou fentes branchiales)
- une queue qui s’étend au-delà de l’ouverture anale (une « queue post anale »)
Nous avons de plus noté que les vertébrés sont des chordés, puisqu’ils ont tous les caractères décrits ci-dessus, mais qu’ils ont aussi un cerveau protégé par un crâne, et une colonne vertébrale. Les chordés non vertébrés incluent deux groupes qui ont des représentants aujourd’hui : les céphalocordés, et les tuniciers. Ces trois groupes monophylétiques forment le groupe couronne des chordés (c’est-à-dire tous les chordés en vie, leur dernière population ancestrale commune, et toutes les espèces descendantes, en vie ou non). Le groupe couronne des vertébrés est à son tour inclus dans les chordés, et les chordés eux-mêmes sont inclus dans un groupe plus large que l’on appelle les deutérostomiens :
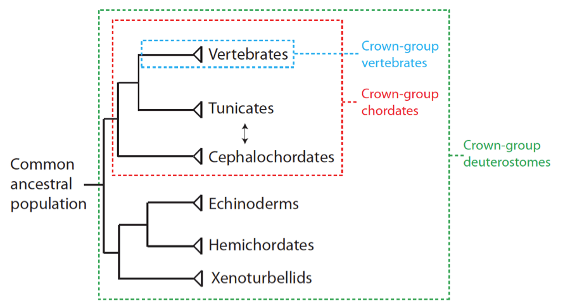
Relations phylogénétiques entre deutérostomes, qui montrent les relations d’inclusion des vertébrés (boîte bleue) avec les chordés (boîte rouge), et des chordés avec les deutérostomes (boîte verte). Tous les vertébrés sont donc des chordés, mais tous les chordés ne sont pas des vertébrés, et ainsi de suite. La flèche indique que la position des tuniciers et des céphalocordés peut en fait être renversée. Ce schéma n’est pas à l’échelle.
La construction du plan d’organisation des vertébrés : le groupe-souche des deutérostomes
Une fois que nous avons localisé les vertébrés dans leur contexte phylogénétique plus large, nous pouvons retrouver certaines innovations clés qui ont conduit à leur plan d’organisation. Comme nous l’avons vu dans le dernier billet, ce processus ne se fonde pas sur la recherche d’ancêtres directs des groupes couronne des vertébrés, mais sur la recherche dans les données fossiles d’organismes qui se sont séparés du lignage des vertébrés à différents moments, emportant avec eux leur ensemble de caractéristiques qui leur étaient propres à ce moment là. En d’autres termes, nous pouvons inférer de nombreuses choses sur le lignage véritable des vertébrés en examinant des groupes souche d’espèces dans les données fossiles.
L’une des difficultés qui se posent à la reconstruction de l’arbre généalogique des vertébrés est le fait que le groupe souche des deutérostomes (et en général les groupes souche des chordés) se composait d’animaux au corps mou, et ne se fossilisent donc pas facilement. En effet, ce que nous savons de ces espèces anciennes et au corps mou nous vient de quelques sites de fossiles clé, où des circonstances fortuites ont conduit à leur fossilisation : des sites comme les schistes de Burgess au Canada et le site fossilifère de Chengjiang en Chine. Sur ces sites, nous voyons des « instantanés » géologiques de la vie de la période cambrienne. Les fossiles de Chengjian capturent une fenêtre d’il y a 525 à 520 millions d’années, et les schistes de Burgess capturent une période un peu plus tardive du cambrien, il y a 505 millions d’années. De plus, la vie florissante que l’on voit sur ces sites (et à ces moments), préservée de façon exceptionnelle, indique que les données fossiles « normales », où les circonstances n’ont pas favorisé la fossilisation de corps mous, sont malheureusement éparses.
Les Vetilucolia sont un groupe d’organismes qui éclairent les origines des deutérostomes ; ils forment un groupe d’organismes resté plutôt mystérieux jusqu’à de récents travaux basés sur de nouveaux fossiles trouvés à Chengjiang ; ceux-ci ont montré qu’il était un groupe souche probable du lignage deutérostome. Les preuves en faveur de l’idée selon laquelle les Vetilucolia sont un groupe souche des deutérostomes se fondent sur ce qui semble être un pharynx et des sillons pharyngés. Ces structures leur auraient permis de rejeter de l’eau de mer avalée et de se nourrir en suspensivores. Le fait qu’un groupe d’espèces ait ces traits – sans aucune autre des caractéristiques qui définissent le groupe couronne des deutérostomes – suggère fortement qu’un premier pas vers le « plan d’organisation des vertébrés » a consisté dans le développement des sillons pharyngés :

La construction du plan d’organisation des vertébrés : le groupe souche des chordés
Le cambrien donne aussi des éléments qui montrent qu’il y aurait eu un groupe souche des chordés en plus du groupe souche des deutérostomes. Le Pikaia gracilens par exemple, trouvé dans les schistes de Burgess, est une espèce dont on a immédiatement noté la ressemblance avec les céphalochordés lorsqu’on l’a découvert dans les années 1990. Une réévaluation récente à grande échelle de Pikaia le situe comme groupe souche des chordés. En effet, Pikaia a un pharynx avec des sillons pharyngés, une notochorde, et une corde dorsale nerveuse vide, mais il ne possède pas d’autres caractères propres aux chordés (tels que la queue post anale). Ceci soutient l’hypothèse selon laquelle le prochain « pas » vers le groupe couronne des vertébrés a été le développement du nerf dorsal et de la notochorde :

Ainsi la prochaine caractéristique à être acquise serait une queue post anale, pour finalement arriver aux caractères qui définissent ce que nous reconnaissons désormais comme un groupe monophylétique largement réussi : le groupe couronne des chordés.
La construction du plan d’organisation des vertébrés : le groupe couronne cambrien des vertébrés
La région de Chengjiang est peut-être surtout célèbre pour la découverte d’espèces cambriennes que l’on pense être de véritables vertébrés, incluant Myllokunmingia Fengjiaoa et Haikouichthys ercaicunensis. On pense que ces espèces sont les exemples connus les plus anciens de poissons sans mâchoire, avec un crâne et une épine dorsale en cartilage. Si ces espèces sont effectivement de véritables vertébrés, elles feraient partie du groupe couronne des vertébrés (bien que, comme nous le verrons dans d’autres billets, ces espèces sont aussi un groupe souche par rapport à des sous-catégories plus tardives de vertébrés avec des caractéristiques en plus, comme la mâchoire) :
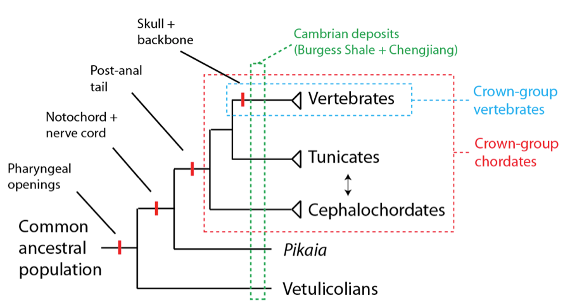
En résumé : des deutérostomes aux vertébrés
Pris ensemble, nous avons vu que bien que le plan d’organisation des vertébrés soit d’abord apparu dans les données fossiles de la période cambrienne (grâce à sa préservation fortuite à Chengjiang), il y a de nombreuses espèces dans le cambrien que l’on peut identifier comme des groupes souche sur le lignage des vertébrés. Ces groupes souche nous montrent que le plan d’organisation des vertébrés a été mis en place au cours du temps étape par étape, et que son « apparition soudaine » dans les données fossiles cambriennes n’est en fait pas soudaine du tout, mais plutôt le résultat final d’un processus dont l’extension s’enracine plus profondément dans le passé.
Dans le prochain billet de cette série, nous suivrons le lignage vertébré vers des temps plus récents, et examinerons la merveilleuse diversité de formes présentes dans le groupe couronne des vertébrés.
Pour en savoir plus
Swalla, B.J. and Smith, A.B. (2008). Deciphering deuterostome phylogeny: molecular, morphological and palaeontological perspectives. Phil. Trans. R. Soc. B 363, doi: 10.1098/rstb.2007.2246.
http://rstb.royalsocietypublishing.org/content/363/1496/1557.long
Morris, S.C. and Caron, J-B. (2012). Pikaia gracilens Walcott, a stem-group chordate from the Middle Cambrian of British Columbia. Biol. Rev. 87; 480–512.
http://www.ncbi.nlm.nih.gov/pubmed/22385518
Ou, Q., et al. (2012). Evidence for gill slits and a pharynx in Cambrian vetulicolians: implications for the early evolution of deuterostomes. BMC Biology 10:81.
http://www.ncbi.nlm.nih.gov/pubmed/23031545
48 Articles pour la série :
- 01-L’évolution expliquée : Introduction
- 02-L’Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l’homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l’humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour


